
Rationale and Objectives
The study was performed to evaluate the effect of magnetic resonance imaging (MRI) contrast agent (super paramagnetic iron oxide [SPIO]) on differentiation and migration of primary murine neural stem cells (NSCs) in comparison to a neural stem cell line (C17.2). Because detection of labeled cells depends on the concentration of SPIO particles per imaging voxel, the study was performed at various concentrations of SPIO particles to determine the concentration that could be used for in vivo detection of small clusters of grafted cells.
Materials and Methods
Murine primary NSCs or C17.2 cells were labeled with different concentrations of SPIO particles (0, 25, 100, and 250 μg Fe/mL) and in vitro assays were performed to assess cell differentiation. In vivo MRI was performed 7 weeks after neonatal transplantation of labeled cells to evaluate the difference in migration capability of the two cell populations.
Results
Both the primary NSCs and the C17.2 cells differentiated to similar number of neurons (Map2ab-positive cells). Similar patterns of engraftment of C17.2 cells were seen in transplanted mice regardless of the SPIO concentration used. In vivo MRI detection of grafted primary and C17.2 cells was only possible when cells were incubated with 100 μg/mL or higher concentration of SPIO. Extensive migration of C17.2 cells throughout the brain was observed, whereas the migration of the primary NSCs was more restricted.
Conclusions
Engraftment of primary NSCs can be detected noninvasively by in vivo MRI, and the presence of SPIO particles do not affect the viability, differentiation, or engraftment pattern of the donor cells.
Stem cells have the ability to differentiate into specific cell types. They have been used in several preclinical models of disease ( ) and are being used in Phase I–III clinical trials ( ). Neural stem cells (NSCs) can differentiate into neuronal or glial cells and express trophic factors to rescue dysfunctional brain tissue ( ). These properties of NSCs provide an opportunity to use them as delivery vehicles for therapeutic molecules into specific regions or into the entire brain ( ). Stem cell lines are generally used for investigation of basic properties of stem cells. One of the commonly used NSC lines, known as C17.2, was originally derived from neonatal mouse cerebellum and immortalized by the introduction of a v-myc oncogene ( ). When transplanted into the developing brain, this cell line consistently results in robust and stable engraftment throughout the brain. Because this line does not usually lead to formation of tumors, it has been extensively used in transplantation biology studies, including treatment of mouse models of lysosomal storage diseases ( ). However, use of neural stem cell lines for treatment of diseases in humans is potentially problematic because of their immortalized nature. An earlier study suggested that the biology of C17.2 cells differs significantly from primary neural stem cells ( ); therefore, it is important to assess the transplantation and migration properties of primary stem cells in experimental models.
Stem cell engraftment is generally determined postmortem by histologic, immunologic, or fluorescence assays. However, to evaluate the efficacy of stem cell migration and survival over time, it is necessary to use noninvasive techniques. Magnetic resonance imaging (MRI) methods have been applied to monitor implanted stem cells by loading the cells with iron oxide particles as a contrast agent ( ). The particles exhibit strong magnetic moments when placed in a magnetic field and create a hypointense (dark) signal on MRI. Based on their size, these agents can be broadly classified into three categories; ultra-small paramagnetic iron oxide particles (USPIO, 10–30 nm), super paramagnetic iron oxide particles (SPIO, 30–150 nm) and micrometer-sized paramagnetic iron oxide particles (0.5–2 μm). The SPIO particles are more commonly used ( ) because they are commercially available and have been used in the clinic for the detection of liver tumors ( ) and have recently been shown to be efficient in the detection of implanted cells in humans ( ).
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Methods
Experimental Animals
Get Radiology Tree app to read full this article<
Isolation of Mouse Neural Precursor Cells
Get Radiology Tree app to read full this article<
Primary Mouse Neural Precursor Cell Culture
Get Radiology Tree app to read full this article<
Cell Culture of C17.2 Cells
Get Radiology Tree app to read full this article<
Labeling of Cells with Iron Oxide Particles
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Differentiation Assay of NSCs
Get Radiology Tree app to read full this article<
Intracranial Cell Implantation
Get Radiology Tree app to read full this article<
MRI Experiments
In vivo imaging
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Ex vivo imaging
Get Radiology Tree app to read full this article<
Image analysis
Get Radiology Tree app to read full this article<
Histology
Get Radiology Tree app to read full this article<
Prussian blue staining for detection of iron
Get Radiology Tree app to read full this article<
β-galactosidase staining
Get Radiology Tree app to read full this article<
GUSB staining
Get Radiology Tree app to read full this article<
CD11b staining
Get Radiology Tree app to read full this article<
Results
Iron Uptake of Cells
Get Radiology Tree app to read full this article<
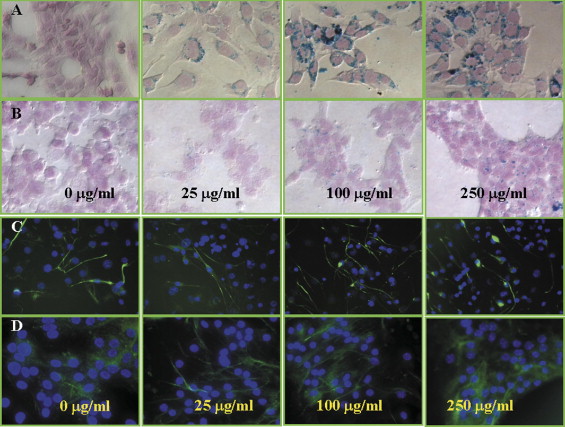
Table 1
Net Iron Concentration per Cell as Measured by Inductively Coupled Plasma Mass Spectroscopy after Incubation of Cells with a Solution of Ferumoxide-poly- d -lysine
Iron Concentration per Cell (pg/cell) Iron Concentration in Incubation Medium (mg/mL) 0 25 100 250 C17.2 <0.15 10.6 ± 1.3 46.5 ± 27.5 152.9 ± 112 ⁎ Primary neural stem cells <0.15 5.5 ± 2.2 4.5 ± 2.1 8.3 ± 9.0 ⁎
Values are reported as mean ± standard deviation.
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Cell Differentiation in the Presence of SPIO Particles
Get Radiology Tree app to read full this article<
Table 2
Percentage of Map2ab (neuronal)-positive C17.2 and Murine Primary Stem Cells
% Map2ab Positive Iron Concentration in Incubation Medium (mg/mL) 0 25 100 250 C17.2 cells 9.08 ± 3.9 10.9 ± 4.7 11.6 ± 7.9 11.3 ± 5.3 Primary neural stem cells 5.4 ± 4.7 1.7 ± 1.9 6.3 ± 5.9 10.1 ± 6.0
C17.2 cells were differentiated for 5 days, whereas primary cells were differentiated for approximately 10 days. Values are reported as the average number of positive cells with respect to the total number of cells ± standard deviaiton (from five random fields and three separate experiments).
Get Radiology Tree app to read full this article<
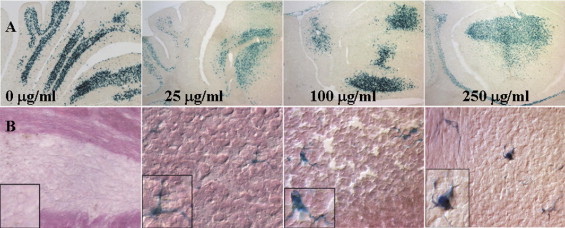
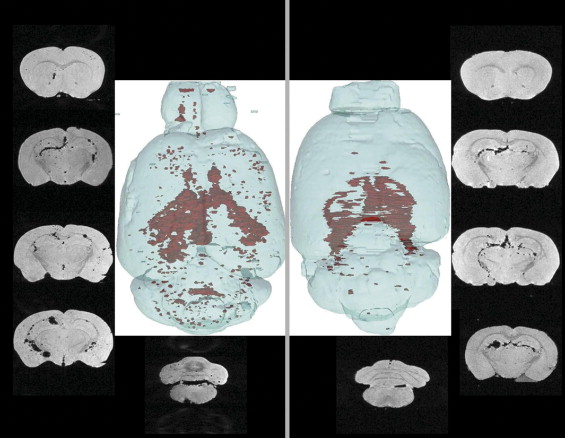
Engraftment Pattern
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
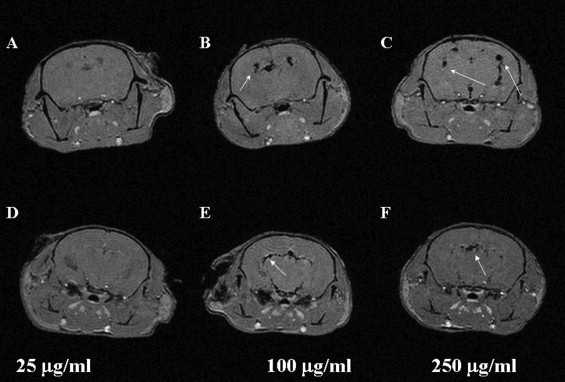
MRI Detection of Labeled C17.2 Cells
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
MRI Detection of Murine Primary Stem Cells
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
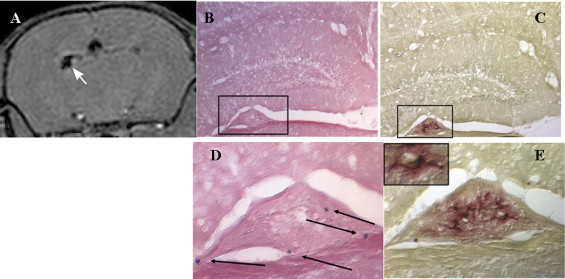
Detection of Iron-positive Grafted Cells and Microglia
Get Radiology Tree app to read full this article<
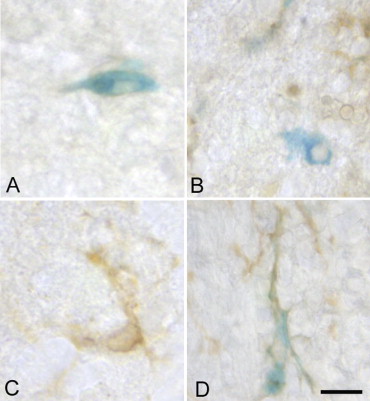
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Discussion
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
Get Radiology Tree app to read full this article<
References
1. Hematti P., Obrtlikova P., Kaufman D.S.: Nonhuman primate embryonic stem cells as a preclinical model for hematopoietic and vascular repair. Exp Hematol 2005; 33: pp. 980-986.
2. Martino G., Pluchino S.: The therapeutic potential of neural stem cells. Nat Rev Neurosci 2006; 7: pp. 395-406.
3. Zhang R.L., Zhang Z.G., Chopp M.: Neurogenesis in the adult ischemic brain: generation, migration, survival, and restorative therapy. Neuroscientist 2005; 11: pp. 408-416.
4. Kuthiala S.S., Lyman G.H., Ballester O.F.: Randomized clinical trials for hematopoietic stem cell transplantation: lessons to be learned from the European experience. Bone Marrow Transplant 2006; 37: pp. 219-221.
5. Nash R.A., Bowen J.D., McSweeney P.A., et. al.: High-dose immunosuppressive therapy and autologous peripheral blood stem cell transplantation for severe multiple sclerosis. Blood 2003; 102: pp. 2364-2372.
6. Okada H., Pollack I.F., Lieberman F., et. al.: Gene therapy of malignant gliomas: a pilot study of vaccination with irradiated autologous glioma and dendritic cells admixed with IL-4 transduced fibroblasts to elicit an immune response. Human Gene Ther 2001; 12: pp. 575-595.
7. Hesdorffer C., Ayello J., Ward M., et. al.: Phase I trial of retroviral-mediated transfer of the human MDR1 gene as marrow chemoprotection in patients undergoing high-dose chemotherapy and autologous stem-cell transplantation. J Clin Oncol 1998; 16: pp. 165-172.
8. Snyder E.Y., Deitcher D.L., Walsh C., et. al.: Multipotent neural cell lines can engraft and participate in development of mouse cerebellum. Cell 1992; 68: pp. 33-51.
9. Snyder E.Y., Taylor R.M., Wolfe J.H.: Neural progenitor cell engraftment corrects lysosomal storage throughout the MPS VII mouse brain. Nature 1995; 374: pp. 367-370.
10. Flax J.D., Aurora S., Yang C., et. al.: Engraftable human neural stem cells respond to developmental cues, replace neurons, and express foreign genes. Nat Biotechnol 1998; 16: pp. 1033-1039.
11. Fricker R.A., Carpenter M.K., Winkler C., et. al.: Site-specific migration and neuronal differentiation of human neural progenitor cells after transplantation in the adult rat brain. J Neurosci 1999; 19: pp. 5990-6005.
12. Ourednik J., Ourednik V., Lynch W.P., et. al.: Neural stem cells display an inherent mechanism for rescuing dysfunctional neurons. Nat Biotechnol 2002; 20: pp. 1103-1110.
13. Wang Y., Alexander O.B., Woodward-Pu Y.M., et. al.: Viral vector strategy for glial cell line-derived neurotrophic factor therapy for stroke. Frontiers Biosci 2006; 11: pp. 1101-1107.
14. Watson D.J., Walton R.M., Magnitsky S., et. al.: Structure-specific patterns of neural stem cell engraftment after transplantation in the adult mouse brain. Human Gene Ther 2006; 17: pp. 693-704.
15. Ryder E.F., Snyder E.Y., Cepko C.L.: Establishment and characterization of multipotent neural cell lines using retrovirus vector-mediated oncogene transfer. J Neurobiol 1990; 21: pp. 356-375.
16. Riess P., Zhang C., Saatman K.E., et. al.: Transplanted neural stem cells survive, differentiate, and improve neurological motor function after experimental traumatic brain injury. Neurosurgery 2002; 51: pp. 1043-1052. discussion 1052–1054
17. Yandava B.D., Billinghurst L.L., Snyder E.Y.: “Global” cell replacement is feasible via neural stem cell transplantation: evidence from the dysmyelinated shiverer mouse brain. Proc Natl Acad Sci U S A 1999; 96: pp. 7029-7034.
18. Park K.I., Teng Y.D., Snyder E.Y.: The injured brain interacts reciprocally with neural stem cells supported by scaffolds to reconstitute lost tissue. Nat Biotechnol 2002; 20: pp. 1111-1117.
19. Lacorazza H.D., Flax J.D., Snyder E.Y., et. al.: Expression of human beta-hexosaminidase alpha-subunit gene (the gene defect of Tay-Sachs disease) in mouse brains upon engraftment of transduced progenitor cells. Nat Med 1996; 2: pp. 424-429.
20. Mi R., Luo Y., Cai J., Limke T.L., et. al.: Immortalized neural stem cells differ from nonimmortalized cortical neurospheres and cerebellar granule cell progenitors. Exp Neurol 2005; 194: pp. 301-319.
21. Bulte J.W., Douglas T., Witwer B., et. al.: Magnetodendrimers allow endosomal magnetic labeling and in vivo tracking of stem cells. Nat Biotechnol 2001; 19: pp. 1141-1147.
22. Modo M., Hoehn M., Bulte J.W.: Cellular MR imaging. Mol Imaging 2005; 4: pp. 143-164.
23. Hoehn M., Kustermann E., Blunk J., et. al.: Monitoring of implanted stem cell migration in vivo: a highly resolved in vivo magnetic resonance imaging investigation of experimental stroke in rat. Proc Natl Acad Sci U S A 2002; 99: pp. 16267-16272.
24. Magnitsky S., Walton R.M., Wolfe J.H., et. al.: Magnetic resonance imaging as a tool for monitoring stem cell migration. Neurodegen Dis 2007; 4: pp. 314-321.
25. Bulte J.W., Arbab A.S., Douglas T., et. al.: Preparation of magnetically labeled cells for cell tracking by magnetic resonance imaging. Methods Enzymol 2004; 386: pp. 275-299.
26. Frank J.A., Anderson S.A., Kalsih H., et. al.: Methods for magnetically labeling stem and other cells for detection by in vivo magnetic resonance imaging. Cytotherapy 2004; 6: pp. 621-625.
27. Schultz J.F., Bell J.D., Goldstein R.M., et. al.: Hepatic tumor imaging using iron oxide MRI: comparison with computed tomography, clinical impact, and cost analysis. Ann Surg Oncol 1999; 6: pp. 691-698.
28. de Vries I.J., Lesterhuis W.J., Barentsz J.O., et. al.: Magnetic resonance tracking of dendritic cells in melanoma patients for monitoring of cellular therapy. Nat Biotechnol 2005; 23: pp. 1407-1413.
29. Magnitsky S., Watson D.J., Walton R.M., et. al.: In vivo and ex vivo MRI detection of localized and disseminated neural stem cell grafts in the mouse brain. NeuroImage 2005; 26: pp. 744-754.
30. Jendelova P., Herynek V., Urdzikova L., et. al.: Magnetic resonance tracking of transplanted bone marrow and embryonic stem cells labeled by iron oxide nanoparticles in rat brain and spinal cord. J Neurosci Res 2004; 76: pp. 232-243.
31. Daldrup-Link H.E., Meier R., Rudelius M., et. al.: In vivo tracking of genetically engineered, anti-HER2/neu directed natural killer cells to HER2/neu positive mammary tumors with magnetic resonance imaging. Eur Radiol 2005; 15: pp. 4-13.
32. Kostura L., Kraitchman D.L., Mackay A.M., et. al.: Feridex labeling of mesenchymal stem cells inhibits chondrogenesis but not adipogenesis or osteogenesis. NMR Biomed 2004; 17: pp. 513-517.
33. Pawelczyk E., Arbab A.S., Elbert Hu S.P., et. al.: Expression of transferrin receptor and ferritin following ferumoxides-protamine sulfate labeling of cells: implications for cellular magnetic resonance imaging. NMR Biomed 2006; 19: pp. 581-592.
34. Arbab A.S., Bashaw L.A., Miller B.R., et. al.: Characterization of biophysical and metabolic properties of cells labeled with superparamagnetic iron oxide nanoparticles and transfection agent for cellular MR imaging. Radiology 2003; 229: pp. 838-846.
35. Walczak P., Kedziorek D.A., Gilad A.A., et. al.: Applicability and limitations of MR tracking of neural stem cells with asymmetric cell division and rapid turnover: the case of the shiverer dysmyelinated mouse brain. Magn Res Med 2007; 58: pp. 261-269.
36. Kyle J.W., Birkenmeier E.H., Gwynn B., et. al.: Correction of murine mucopolysaccharidosis VII by a human beta-glucuronidase transgene. Proc Natl Acad Sci U S A 1990; 87: pp. 3914-3918.
37. Frank J.A., Miller B.R., Arbab A.S., et. al.: Clinically applicable labeling of mammalian and stem cells by combining superparamagnetic iron oxides and transfection agents. Radiology 2003; 228: pp. 480-487.
38. Wolfe J.H., Sands M.S.: Murine mucopolysaccharidosis Type VII: a model system for somatic gene therapy of the central nervous system.Lowenstein P.R.Enquist L.W.Protocols for gene transfer in neuroscience: towards gene therapy of neurological disorders.1996.John Wiley & Sons LtdNew York:pp. 263-274.
39. Jendelová P., Herynek V., DeCroos J., et. al.: Imaging the fate of implanted bone marrow stromal cells labeled with superparamagnetic nanoparticles. Magn Res Med 2003; 50: pp. 767-776.
40. Arbab A.S., Jordan E.K., Wilson L.B., et. al.: In vivo trafficking and targeted delivery of magnetically labeled stem cells. Human Gene Ther 2004; 15: pp. 351-360.
41. Arbab A.S., Yocum G.T., Rad A.M., et. al.: Labeling of cells with ferumoxides-protamine sulfate complexes does not inhibit function or differentiation capacity of hematopoietic or mesenchymal stem cells. NMR Biomed 2005; 18: pp. 553-559.
42. Bulte J.W.M., Kraitchman D.L., Mackay A.M., et. al.: Chondrogenic differentiation of mesenchymal stem cells is inhibited after magnetic labeling with ferumoxides. Blood 2004; 104: pp. 3410-3413.
43. Brekke C., Morgan S.C., Lowe A.S., et. al.: The in vitro effects of a bimodal contrast agent on cellular functions and relaxometry. NMR Biomed 2007; 20: pp. 77-89.
44. Modo M., Mellodew K., Cash D., et. al.: Mapping transplanted stem cell migration after a stroke: a serial, in vivo magnetic resonance imaging study. Neuroimage 2004; 21: pp. 311-317.
45. Jackson P.K.: Developmental neurobiology: a destructive switch for neurons. Nature 2006; 442: pp. 365-366.
46. Heuer G.G., Passini M.A., Jiang K., et. al.: Selective neurodegeneration in murine mucopolysaccharidosis VII is progressive and reversible. Ann Neurol 2002; 52: pp. 762-770.
47. Tamm C., Duckworth J., Hermanson O., et. al.: High susceptibility of neural stem cells to methylmercury toxicity: effects on cell survival and neuronal differentiation. J Neurochem 2006; 97: pp. 69-78.
48. Stroh A., Faber C., Neuberger T., et. al.: In vivo detection limits of magnetically labeled embryonic stem cells in the rat brain using high-field (17.6 T) magnetic resonance imaging. Neuroimage 2005; 24: pp. 635-645.
49. Foster-Gareau P., Heyn C., Alejski A., et. al.: Imaging single mammalian cells with a 1.5 T clinical MRI scanner. Magn Res Med 2003; 49: pp. 968-971.
50. Daldrup-Link H.E., Rudelius M., Metz S., et. al.: Cell tracking with gadophrin-2: a bifunctional contrast agent for MR imaging, optical imaging, and fluorescence microscopy. Eur J Nucl Med Mol Imaging 2004; 31: pp. 1312-1321.
51. Shah K., Weissleder R.: Molecular optical imaging: applications leading to the development of present day therapeutics. NeuroRx 2005; 2: pp. 215-225.
52. Acton P.D., Zhou R.: Imaging reporter genes for cell tracking with PET and SPECT. Q J Nucl Med Mol Imaging 2005; 49: pp. 349-360.
53. Adonai N., Nguyen K.N., Walsh J., et. al.: Ex vivo cell labeling with 64Cu-pyruvaldehyde-bis(N4-methylthiosemicarbazone) for imaging cell trafficking in mice with positron-emission tomography. Proc Natl Acad Sci U S A 2002; 99: pp. 3030-3035.
54. Bogdanov A.A., Simonova M., Weissleder R.: Engineering membrane proteins for nuclear medicine: applications for gene therapy and cell tracking. Q J Nucl Med 2000; 44: pp. 224-235.
55. Herschman H.R.: PET reporter genes for noninvasive imaging of gene therapy, cell tracking and transgenic analysis. Crit Rev Oncol Hematol 2004; 51: pp. 191-204.
56. Shah K., Jacobs A., Breakefield X.O., et. al.: Molecular imaging of gene therapy for cancer. Gene Ther 2004; 11: pp. 1175-1187.
57. Mayer-Kuckuk P., Gade T.P.F., Buchanan I.M., et. al.: High-resolution imaging of bone precursor cells within the intact bone marrow cavity of living mice. Mol Ther 2005; 12: pp. 33-41.
58. Sumner J.P., Conroy R., Shapiro E.M., et. al.: Delivery of fluorescent probes using iron oxide particles as carriers enables in-vivo labeling of migrating neural precursors for magnetic resonance imaging and optical imaging. J Biomed Optics 2007; 12: pp. 051504.
59. Wiart M., Davoust N., Pialat J.B., et. al.: MRI monitoring of neuroinflammation in mouse focal ischemia. Stroke 2007; 38: pp. 131-137.
60. Evgenov N.V., Medarova Z., Pratt J., et. al.: In vivo imaging of immune rejection in transplanted pancreatic islets. Diabetes 2006; 55: pp. 2419-2428.